
胚泡的植入过程
2023-05-05 14:47:47 点击数:当胚胎发育到胚泡期并脱去透明带后,在同步发育的、对胚胎已具备容受性的子宫内膜上开始着床,这个过程分为定位、黏附、植入三个顺序发展阶段。见图4-10。从黏附到植入的过程大约从受精后5~6天开始,在第11~12天完成。
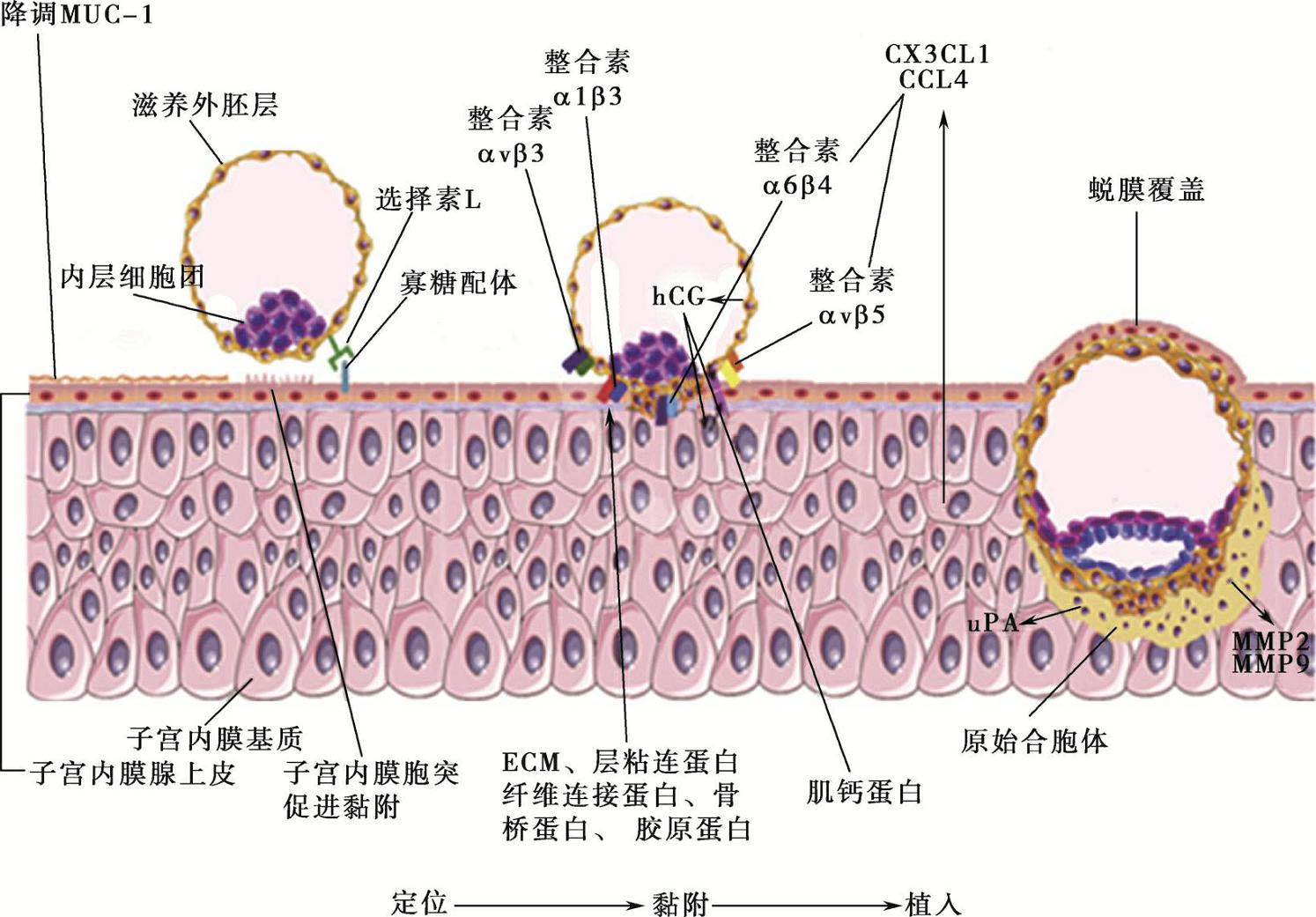
图4-10 胚泡的定位、黏附和植入过程及主要相关分子
受生态、生理和遗传等因素的影响,人类和哺乳动物的胚泡植入方式各不相同。根据胚泡在宫腔的位置和滋养层与子宫内膜接触的程度,将植入分为三种形式:①中央或表面植入型:这种植入方式是胚泡体积长大后充满宫腔一段空间,滋养层大面积与子宫内膜接触,但胚泡不侵入子宫内膜内部。食肉类和部分灵长类动物属于这种类型。②完全侵入植入型:胚泡附着内膜之后侵蚀上皮细胞,然后穿过基底膜钻入内膜基质中。人类、蝙蝠、刺猬和灵长类中的狭鼻猴、猩猩属于这种类型。③偏心植入型:此类胚泡体积较小而宫腔相对较大,胚泡嵌入宫腔一侧上皮陷凹之中,然后上皮消融,胚泡被包埋在基质中。啮齿类多属于此种类型。
(一)定位
受精卵到达宫腔后有1~2天的游离,在黏附之前首先在某个位置靠近内膜,这个过程叫定位(apposition)。定位之后开始进一步黏附直至植入。一般而言,人胚胎在子宫内膜的着床位置大多在子宫后壁或侧壁中部。胚胎选择着床位点的机制尚不清楚。胚胎与子宫内膜之间的相互识别可能是胚胎定位的根本,这种识别主要通过糖复合物与配体的结合介导。
研究表明,在围着床期的子宫内膜和胚泡上存在着大量的糖复合物及配体。糖复合物在子宫内膜的表达除有规律性改变外,还有区域性差别。如兔子宫内膜着床期扁豆凝集素(LCA)、麦芽凝集素(WGA)结合糖蛋白出现阶段性变化,它们的出现与胚泡进入子宫内的时间一致,着床期达顶峰,着床后下降;组化研究表明这种糖复合物主要表达在着床侧;另外,它们的表达量还与胚泡是否存在有关。这说明子宫内膜和胚胎某些特异分子的定点表达与胚泡的定位有关。
最初游离的胚胎靠近子宫内膜可能与两者表面的电荷有关。研究发现,由于子宫内膜上糖基的变化,围着床期子宫内膜的负电荷减少。这增加了内膜对胚胎的吸引力,有利于胚胎靠近内膜。
胚胎具有明显的极性,一般认为人的胚胎是以靠近内细胞团的极端滋养层细胞与子宫内膜相接触,但目前尚无定论。
(二)黏附
在胚泡定位后,开始黏附(adhesion)于子宫内膜,子宫内膜和胚泡之间立即出现微绒毛交错现象,滋养层细胞和子宫内膜上皮细胞间形成桥粒和连接复合体等专门附着的结构。扫描电镜观察发现,胚泡黏附在带胞饮突的上皮细胞上。早期胚泡黏附的第一个形态学标志是顶端连接复合体的形成,它位于上皮细胞的顶端侧面交界处,这使胚泡可以足够稳定地穿过相邻的上皮细胞。
黏附过程主要是通过胚胎和内膜表面表达的多种黏附分子及其配体结合介导的。黏附分子包括整合素、E-选择素、trophinin、细胞间黏附分子-1(ICAM-1)、CD44等,在着床期子宫内膜上大量表达。整合素属于跨膜蛋白类黏附分子,它们的配体包括层粘连蛋白(laminin)、纤连蛋白(fibronectin,Fn)、玻连蛋白、胶原蛋白等。整合素介导细胞与细胞、细胞与细胞外基质间的相互作用,促进细胞的迁移、分化和结构变化,整合素-αvβ3、α1β1等周期性地表达在子宫内膜上皮细胞上,在着床期达到峰值。组化研究发现着床部位的Fn远高于非着床部位,有学者形象地把Fn称为胚胎和子宫内膜间黏附的“分子胶”,Fn和配体的结合介导了胚胎在子宫内膜上的黏附过程。研究表明,着床期子宫内膜上整合素的表达缺失会导致不孕。
(三)植入
1.胚泡植入
胚泡附着于子宫内膜表面后,很快侵入(penetration)子宫内膜。滋养层穿入子宫内膜的过程称植入(implantation)。胚泡的植入阶段包括滋养细胞与细胞外基质(extra-cellular matrix,ECM)的黏附、基质降解、内膜蜕膜化、细胞滋养细胞移行等变化。胚泡穿过子宫内膜上皮层时可以使上皮间的桥粒和顶端复合体解离。胚泡植入内膜的形式因种属的不同而不同,人类的胚胎植入属于完全侵入型。
胚泡植入内膜有三种形式:①融合型侵入:胚泡和子宫内膜接触后,滋养层和内膜上皮细胞顶部发生融合,接触面细胞膜消失,上皮细胞失去原有的特性,融合细胞穿过基底膜进入基质。②移位型侵入:简单地讲就是子宫内膜上皮细胞移位让胚泡侵入。滋养细胞入侵后产生溶酶体酶和蛋白水解酶,使内膜上皮细胞变性坏死,最终脱落,从而使滋养细胞经过基底膜钻入基质。而滋养细胞则可吞噬部分脱落的上皮细胞及其碎片。③闯入型侵入:合体滋养细胞的突起插入上皮细胞相邻之间的缝隙,插入部分扩张,一直延伸到基底膜,穿过基底膜挤入基质内。两类细胞均保持完整。
滋养细胞接触子宫内膜后分泌蛋白水解酶,如MMPs,其中MMP-2、MMP-9等与着床密切相关,可消化与其接触的子宫内膜组织,并降解ECM,使内膜形成缺口,胚泡则通过被消化的缺口逐渐植入子宫内膜功能层。
植入使子宫内膜基质成分发生重建,这主要是蜕膜化过程中的基质成分更新所致。在着床前,ECM主要由纤维胶原Ⅰ和Ⅲ、胶原Ⅴ和Ⅵ、Fn等组成。蜕膜化过程中,子宫内膜基质重新构建,胶原基质纤维性下降、消失,蜕膜细胞产生新的基质成分,如层粘连蛋白、Ⅵ型胶原、entactin等,MMPs在此过程中发挥重要作用。在植入过程中,滋养层细胞迅速增殖分化成内、外两层,内层为细胞滋养层细胞(cytotrophoblast),外层为合体滋养层细胞(syncytiotrophoblast),合体细胞与母血直接接触。胚泡完全植入子宫内膜后,缺口由周围增生的上皮细胞修复,植入过程完成。植入过程持续4天左右。经过植入,原来游离的胚胎埋入子宫内膜中,从而取得母体的营养,建立起母子间结构上的联系。
2.子宫内膜蜕膜化
胚泡植入时,在孕酮的作用下,子宫内膜腺体增大、弯曲,腺体中含有大量黏液及糖原。内膜血管充血,结缔组织细胞肥大,这一系列变化称为蜕膜化(decidual response)。此时的子宫内膜称为蜕膜。电镜观察发现,人类蜕膜中有三类不同的细胞,一种是小细胞,属于未分化细胞,胞质较少;另一种细胞中等大小,胞质内富含糖原和脂类物质;第三种细胞呈斑点状,核糖体和线粒体丰富,核小,核胞膜致密。蜕膜细胞间可见PAS反应阳性的沉积物。
3.胚泡植入时母胎界面的分子对话
胚泡植入是一个精细调节的过程,该过程受母胎界面上表达的大量分子调控,这些分子包括激素、糖复合物、细胞因子、生长因子、黏附分子、蛋白水解酶等。在着床局部母胎界面微环境表达的、参与调节着床的分子很多,研究较多的细胞因子有白血病抑制因子(LIF)、IL-1系统、TGF-B、IGF、EGF、HP-EGF等,黏附分子包括整合素、E-选择素、ICAM-1及其ECM等,此外还有前列腺素、环氧化酶-2等。这些分子在围着床期母胎界面上的表达发生特异性变化。这些分子形成一个复杂的分子网络,在着床过程中进行分子对话(molecules cross-talk),从而激发着床过程。见图4-11。尽管目前已经发现许许多多着床相关分子,但每年的研究仍然在发现新的相关分子,所以,目前尚没有完全解开着床过程的分子调节机制。近几年人们利用基因芯片技术、蛋白组学技术、代谢组学技术筛选到更多的着床相关分子,结合生物信息学方法挖掘分析,找出了一些调节着床的关键信号网络,这为进一步揭示着床机制提供了更好的前景。
- 上一篇:图解子宫内膜容受性建立
- 下一篇:二、捐精会存在相应的伦理问题吗?
