
胎盘分泌的主要激素
2023-05-05 14:52:25 点击数:(一)类固醇激素
类固醇激素(steroid hormones)主要由肾上腺和性腺分泌的激素组成,包括糖皮质激素、盐皮质激素、孕激素、雌激素、雄激素。1,25-二羟基维生素D 3 虽然不是由肾上腺和性腺分泌的,但也属于类固醇激素。胆固醇是类固醇激素的前体,在类固醇激素的结构中都有一个环戊烷多氢菲的基本结构(图5-1)。妊娠初期,孕激素和雌激素主要来源于卵巢黄体,妊娠第8周左右起,胎盘分泌的孕激素和雌激素逐渐增多,于妊娠第10周左右,胎盘完全取代黄体合成孕激素和雌激素的功能,黄体出现萎缩。胎盘虽然不能从头合成糖皮质激素,但存在糖皮质激素的转化酶。为了更好地理解胎盘类固醇激素的合成,本节将首先介绍性腺和肾上腺类固醇激素的经典合成途径。
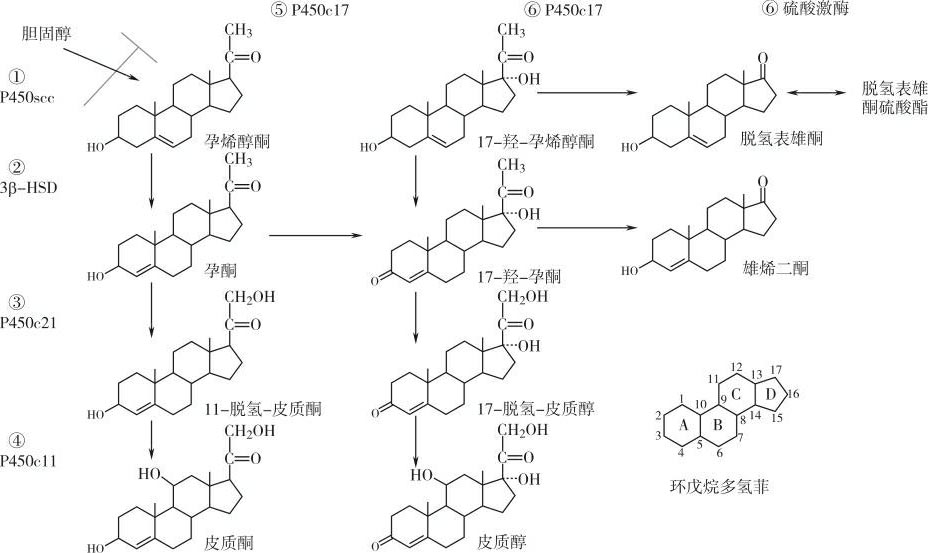
图5-1 类固醇激素的基本结构和合成途径
性腺和肾上腺
卵巢、睾丸和肾上腺的类固醇激素合成的基本途径相似(图5-1),这些腺体都可以利用胆固醇从头合成类固醇激素。它们既可以从血液中的低密度脂蛋白(LDL)摄取胆固醇,又可以从醋酸盐从头合成胆固醇。参与类固醇激素合成的酶体系主要包括3β-羟基类固醇脱氢酶(3β-hydroxysteroid dehydrogenase,3β-HSD)、芳香化酶(aromatase)和四种细胞色素P-450酶,但不同组织的类固醇激素的合成酶体系存在差别,这些差别决定着不同腺体所合成的类固醇激素的不同。类固醇激素合成的第一步反应是胆固醇在细胞色素P-450侧链裂解酶(P-450 scc )的催化作用下,将其C 20 和C 22 之间的碳链断裂,形成孕烯醇酮(pregnenolone),继而孕烯醇酮在滑面内质网酶3β-HSD的催化下,将C 3 位的羧基脱氢氧化为酮基,成为孕酮,又称为孕激素(progesterone)。孕烯醇酮和孕酮均可在另一种滑面内质网细胞色素酶17α-羟化酶(P-450 c17 )的催化下,转化为脱氢表氢酮(dehyroepiandrosterone,DHEA)或雄烯二酮(androstenedione),后两者均可以在17β-羟基类固醇脱氢酶(17β-HSD)的催化下继续转化为睾丸酮(testosterone)或雄烯二醇(androstenediol)。睾丸酮和雄烯二醇则在芳香化酶的催化下分别转化为雌二醇(estradiol)或雌酮(estrone)。孕酮还可在另外两种细胞色素酶的催化下转化为皮质酮(corticosterone)、皮质醇(cortisol)或醛固酮(aldosterone),这两种细胞色素P-450酶分别是位于线粒体的21-羟化酶(P-450 c21 )和位于滑面内质网的11β-羟化酶(P-450 c11 )。肾上腺皮质球状带、束状带和网状带合成的类固醇激素分别以盐皮质激素(醛固酮)、糖皮质激素(皮质醇、皮质酮)和性激素(脱氢表雄酮)为主;睾丸以合成睾丸酮为主;卵巢则以合成雌激素和孕激素为主。
胎盘
胎盘合成的类固醇激素主要为孕烯醇酮、孕酮、雌酮、雌二醇和雌三醇(estriol)。不同于其他类固醇激素的合成组织,胎盘不能利用醋酸盐从头合成胆固醇,只能从血液LDL中摄取胆固醇。所以胎盘合体滋养层细胞膜表面LDL受体成为胎盘类固醇激素合成的限速步骤之一。胎盘表达大量的P-450 scc 和3β-HSD,P-450 scc 可以将胆固醇转化为孕烯醇酮,后者再由3β-HSD转化为孕酮。虽然低等动物胎盘表达的P-450 c17 可以进一步沿着上述合成途径进一步合成雌激素,但是灵长类胎盘缺乏P-450 c17 ,因此不能将孕烯醇酮和孕酮转化为脱氢表雄酮和雄烯二酮,因此灵长类胎盘也就不能利用孕激素合成雌激素。由于灵长类胎盘表达大量的芳香化酶,所以可以利用胎儿和母体肾上腺来源的脱氢表雄酮和雄烯二酮合成雌激素。此外,胎盘还具有很强的再生和失活糖皮质激素的能力,再生和失活糖皮质激素的能力分别依赖于主要在胎膜和胎盘表达的11β-羟基类固醇脱氢酶Ⅰ和Ⅱ型。以下将着重介绍胎盘的孕激素和雌激素及胎盘对糖皮质激素的代谢。
1.孕激素
妊娠初期,母体血液的孕激素来源于卵巢黄体,妊娠第8周以后,则主要来源于胎盘。在妊娠8~10周后切除黄体或双侧卵巢,既不影响妊娠过程,也不影响尿中孕激素代谢产物的排出量,说明此时胎盘的孕激素合成功能已经完全取代了卵巢黄体的孕激素合成功能。
母体血孕激素的水平随妊娠期逐渐增高,妊娠晚期也继续升高,这一点不同与低等动物妊娠晚期出现的孕激素水平的下降,即孕激素撤退。在人类,孕期末母体血液孕激素的浓度可以达到60~100ng/ml。
(1)孕激素的合成:
排卵后,残余的卵泡壁内陷,大量新生血管长入,转变为含有粒黄体细胞与膜黄体细胞的黄体。LH通过cAMP-蛋白激酶A系统,促使黄体细胞分泌大量的孕激素与雌激素。子宫内膜在雌激素作用的基础上,受孕激素的刺激进入分泌期,表现为内膜细胞体积增大,糖原含量增加,腺管由直变曲,分泌含糖原的黏液,为受孕作准备。若不受孕,在雌激素和孕激素的负反馈的作用下,垂体FSH和LH的分泌减少,黄体逐渐退化,雌激素和孕激素分泌减少,出现子宫内膜脱落与出血(月经);若受孕,则在受精后第六天左右,胚泡滋养层细胞开始分泌绒毛膜促性腺激素(hCG),hCG具有LH样作用,可以替代垂体LH的作用,继续维持卵巢的黄体功能,使其变为妊娠黄体,并继续分泌孕激素和雌激素。这也是母体和胎儿在妊娠期激素合成过程中的最早合作。但在妊娠第8周时,胎盘合体滋养层细胞开始分泌孕激素,在第10周其合成类固醇激素的功能逐渐成熟,并取代黄体的功能,胎盘成为孕激素和雌激素合成的主要场所。如果此时摘除黄体或卵巢,尿液孕激素的代谢产物水平不受影响。
人类胎盘合成孕激素的功能很强,孕激素的合成和分泌量随孕期逐渐增加(图5-2),此合成特点有别于低等动物妊娠后期出现孕激素水平下降的现象。妊娠后期,胎盘每天分泌的孕激素量可达250~600mg,多胎妊娠时,甚至可以超过600mg。但不像其他合成类固醇激素的腺体,胎盘不能利用醋酸盐从头合成胆固醇,因此只能从母体血液中的LDL摄取胆固醇。如上所述,在由胆固醇合成孕激素的过程中需要P-450 scc 和3β-HSD两种酶的催化作用,在肾上腺皮质,这两种酶都受促肾上腺皮质激素(ACTH)的调控。虽然胎盘的这两种酶的调节目前还不清楚,但胎盘含有大量高活性的P-450 scc 和3β-HSD,在它们的催化下胆固醇极易转化为孕烯醇酮和孕酮。所以,由P-450 scc 和3β-HSD催化的两步反应并不是胎盘孕激素合成的限速步骤,而从LDL中摄取胆固醇则成为胎盘孕激素合成的限速步骤。
1)胆固醇的摄取:
结扎脐带血管、死胎或无头畸形胎儿,胎盘雌激素的合成锐减,但孕激素的合成则基本不受影响,说明胎儿不参与胎盘孕激素的合成。人类胎盘合体滋养层细胞的胆固醇主要来源于母体血液的LDL。合体滋养层细胞对LDL具有很高的摄取能力,其摄取能力依赖于LDL受体的表达,妊娠第六周即有LDL受体的表达。合体滋养层细胞对LDL的水解包括胆固醇酯酶对酯化胆固醇的水解和对LDL载脂蛋白的水解。由酯化胆固醇和载脂蛋白水解产生的必需氨基酸和脂肪酸,可以作为营养物质转运给胎儿。因此,合体滋养层细胞对LDL的水解既为合成类固醇激素提供了前体,又为胎儿提供了营养物质。有一种罕见的家族遗传性低脂蛋白疾病(hypobetalipidemia),患者由于LDL水平降低,体内胆固醇的合成代偿性增加。此类病人妊娠虽然基本正常,但由于可利用的LDL减少,胎盘合成孕激素和雌激素的能力降低,病人血液孕激素和雌激素的水平很低。
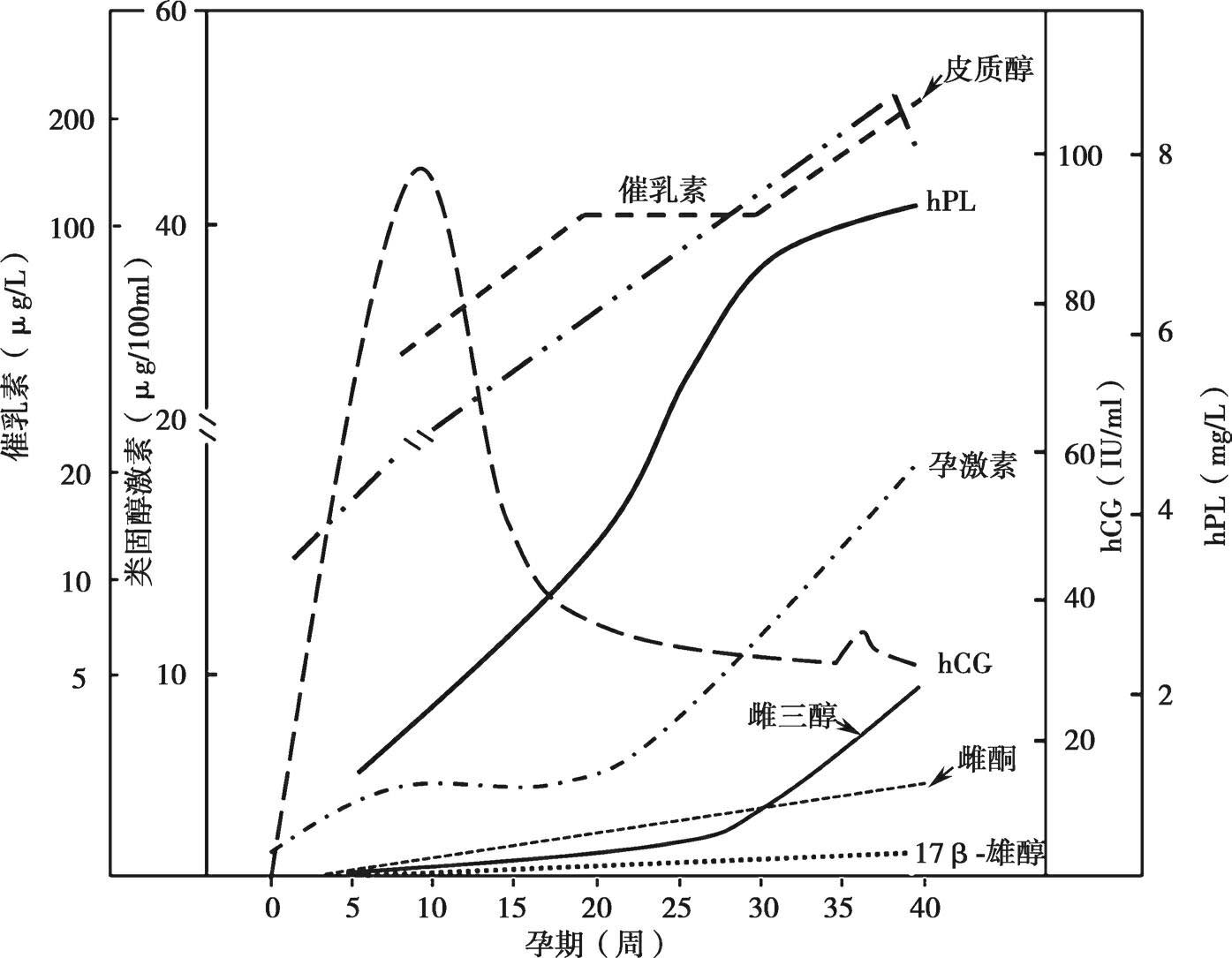
图5-2 人类妊娠期母体血浆激素变化曲线
hCG:人类绒毛膜促性腺激素;hPL:人类胎盘生乳素
2)胆固醇侧链裂解酶:
胎盘细胞由LDL摄取的胆固醇首先在线粒体内由胆固醇侧链裂解酶(P-450 scc )将其转化合体滋养层细胞的微粒体和线粒体内,具有脱氢酶和Δ 5-4 异构酶的双重催化活性:它首先将孕烯醇酮C-3上的羟基脱氢氧化为酮基,再将C-5—C-6之间的双键经Δ 5-4 异构酶的作用,移至C-4—C-5之间成为孕酮(见图5-1)。
体内存在两型3β-HSD:Ⅰ型为孕烯醇酮。P-450 scc 是一种细胞色素酶,其基因位于第15号染色体上,它首先催化胆固醇结构C-22和C-20位的羟基化,然后裂解它们之间的碳链,形成孕烯醇酮。缺乏P-450 scc 的遗传性肾上腺增生症病人,由于 P - 450 scc 基因突变,病人胎盘孕烯醇酮生成减少。
3)3β-羟基类固醇脱氢酶:
妊娠期间,胎盘表达大量的3β-HSD。胎盘3β-HSD除了催化孕烯醇酮向孕酮转化外,还催化脱氢表雄酮向雄烯二酮的转化(图5-3)。3β-HSD存在于胎盘,主要与胎盘孕激素的合成有关;Ⅱ型3β-HSD存在于肾上腺和性腺,为糖皮质激素、盐皮质激素和性激素的合成提供孕酮。Ⅰ型3β-HSD和Ⅱ型3β-HSD是不同基因的产物,Ⅰ型 3β - HSD 基因位于一号染色体上,所表达的Ⅰ型3β-HSD含372个氨基酸残基。当辅酶NAD水平较高时,Ⅰ型3β-HSD表现为氧化酶的活性;当辅酶NADH水平较高时,则表现为还原酶的活性。两型 3β - HSD 基因都是由四个外显子和三个内含子组成,基因长度都为7.8kb,由其转录的mRNA为1.7kb。胎盘 3β - HSD 基因与肾上腺 3β - HSD 基因有多个碱基序列的差别,导致胎盘和肾上腺 3β - HSD 基因的翻译产物有12个氨基酸残基的差别。尽管如此,两型3β-HSD酶结构仍然有90%以上的同源性。肾上腺的Ⅱ型3β-HSD对孕烯醇酮的亲和力低于胎盘Ⅰ型3β-HSD约10倍。根据Ⅰ、Ⅱ型3β-HSD的功能不同,可以设计针对Ⅰ型3β-HSD的特异性抑制剂,特异性阻断妊娠早期孕激素的合成,以达到终止早期妊娠的目的。
(2)孕激素的合成调节:
卵巢黄体孕激素的合成受多种激素的调节,但最为重要的为LH和hCG,它们通过第二信使cAMP促进黄体孕激素的合成和释放。另外,FSH、催乳素和前列腺素等对黄体孕激素的合成也有一定的调节作用。这些激素作用的靶酶为3β-HSD。
胎盘孕激素的合成调节与卵巢各有异同,胎盘hCG为垂体LH的对应激素。过去有文献报道hCG同样也促进胎盘孕激素的合成和释放,但最近的实验均否定了hCG对胎盘孕激素合成的调节作用。为什么hCG调节黄体孕激素的合成而不影响胎盘孕激素的合成目前尚不清楚。虽然hCG对胎盘孕激素的合成没有影响,但生长激素、胰岛素、胰岛素样生长因子-Ⅰ、ACTH、肾上腺素能β受体激动剂和第二信使cAMP类似物可以通过促进P-450 scc 和3β-HSD表达增加胎盘孕激素的合成。PKC激活剂则主要通过促进3β-HSD的表达促进胎盘孕激素的合成。应用芳香化酶抑制剂4-羟雄烯二酮和雌激素受体拮抗剂MER-25可以导致胎盘孕酮的合成减少,提示雌激素促进胎盘孕激素的合成。
(3)孕激素的作用:
孕激素由胎盘分泌入血液后,多数与血液中的转运蛋白(皮质类固醇结合蛋白,CBG)结合运输。
孕激素的作用是通过孕激素受体(progesterone receptor,PR)介导的。PR与其他类固醇激素、甲状腺素、维生素D和视紫红质(retinoids)受体一样,都属于细胞内受体超家族,激活后作为转录因子发挥作用。当孕激素与PR羧基末端的孕激素结合位点结合后,孕激素-受体复合物便与靶基因启动子的孕激素反应元件(progesterone response element,PRE)相结合,调节靶基因的转录。孕激素受体存在PRA和PRB两种亚型,PRA和PRB是同一基因不同转录起始位点的产物,PRA比PRB缺少了一个转录激活区域,虽然两者都能与孕激素结合,但PRA转录激活功能远低于PRB。PRB主要激活靶基因,而PRA为PRB的抑制子。当PRA的表达高于PRB的表达时,孕激素的作用减弱。妊娠晚期PRA/PRB比值上升,导致PRB舒张子宫平滑肌的作用减弱,这样可以在高孕激素环境下削弱孕激素的子宫静息作用,此即所谓的孕激素功能性撤退,不同于低等动物的孕激素合成减少的撤退形式。
孕激素受体存在于人类子宫内膜和腺体的上皮细胞、子宫基质细胞、子宫肌层。排卵前的卵泡颗粒细胞、黄体化的颗粒细胞和黄体也表达孕体激素受体。另外,乳房、中枢的下丘脑腹内侧和视前区、垂体、血管内皮和成骨细胞也都有孕激素受体的表达。雌激素促进PR的表达,孕激素则反馈抑制PR的表达。
1)孕激素对子宫肌的舒张作用:
妊娠时子宫平滑肌的安静状态需要孕激素的维持。孕激素防止子宫平滑肌收缩的机制包括调节细胞内钙离子的水平和调节前列腺素、催产素和舒缓素的作用等。
①对细胞内钙离子水平的调节:钙离子是肌肉兴奋收缩耦联的关键。钙离子转运蛋白calbindin-D9k通过转运钙离子增加子宫平滑肌细胞内钙离子的水平。妊娠期随着孕激素水平的增加,子宫平滑肌的calbindin-D9k表达受到抑制,细胞内钙离子水平降低,子宫平滑肌的兴奋性也随之降低。孕激素受体拮抗剂RU38486可以阻断calbindin-D9k表达的下降,说明此作用是由孕激素受体介导的。
降钙素是调节血钙平衡的重要激素,它通过抑制溶骨过程和肾脏对钙离子的重吸收降低血钙水平,从而使细胞内钙离子水平降低,抑制子宫平滑肌的兴奋-收缩耦联。虽然降钙素主要是由甲状腺滤泡旁细胞合成和分泌的,但子宫内膜腺体细胞在妊娠早期即有降钙素的表达,孕激素对子宫内膜降钙素的表达具有促进作用。
②对前列腺素作用的调节:前列腺素F 2α 和前列腺素E为刺激子宫收缩和宫颈成熟的激素。孕激素通过多种途径抑制前列腺素F 2α 和前列腺素E素对子宫的作用。妊娠期间,高水平的孕激素抑制前列腺素的合成、促进前列腺素降解酶PGDH的表达,从而使子宫内膜前列腺素F 2α 和前列腺素E的水平降低,有利于子宫静息状态的维持。妊娠晚期,许多种属动物(灵长类除外)都出现孕激素向雌激素的转化增加,在孕激素水平降低的同时,雌激素水平升高,同时前列腺素的作用加强,导致分娩的启动。根据此原理,临床上经常应用孕激素受体拮抗剂RU38486进行引产。
③对肽类激素作用的调节:分娩一旦启动后,催产素将强烈收缩子宫促进分娩。催产素主要是由垂体后叶所分泌的。但近年来发现,子宫内组织分泌的催产素也在分娩当中发挥重要的作用。孕激素抑制子宫催产素的表达,PGF 2α 则促进其表达。孕激素还抑制子宫平滑肌血管紧张素-Ⅱ和心房肽受体的表达,血管紧张素-Ⅱ和心房肽都是收缩子宫的激素。舒缓素为妊娠期重要的子宫舒张激素,它不仅抑制子宫平滑肌的自发性收缩,而且还抑制由前列腺素导致的子宫平滑肌的收缩。孕激素可以通过促进黄体、蜕膜和胎盘等部位的舒缓素表达,舒张子宫平滑肌。
④对肾上腺素能系统的调节:妊娠期子宫平滑肌的肾上腺素能β受体系统通过第二信使cAMP参与维持子宫的静息。实验发现,孕激素促进妊娠晚期大鼠子宫平滑肌肾上腺素能β受体的表达。
⑤降低平滑肌细胞膜的电兴奋性:实验发现,孕激素能使一些动物的子宫平滑肌细胞膜发生超极化,对刺激的阈值升高,从而降低子宫平滑肌的兴奋性。
2)孕激素对乳房的作用:
青春期乳腺的发育主要依靠雌激素和孕激素的作用,前者促进乳腺导管的发育,后者促进乳腺小叶的发育,但这时乳腺尚不能产生乳汁。妊娠时,雌激素和孕激素水平进一步升高,加上催乳素水平的升高,导致乳腺进一步增生,为分娩后排乳作好准备。缺乏孕激素受体的小鼠,雌激素和催乳素虽然可以促进乳房小叶导管的增生,但腺泡的发育发生障碍。月经周期中,随着孕激素的周期性变化,乳房腺体细胞的分裂也呈周期性变化,其DNA的合成高峰与黄体晚期孕激素的合成高峰相吻合。孕激素的促进乳腺小叶增生作用可能是通过调节生长因子及其受体的表达实现的。EGF、TGF-α、胰岛素等生长因子促进乳腺细胞的生长分化,而TGF-β则抑制乳腺细胞的生长。孕激素促进乳腺TGF-α、EGF及其受体、胰岛素受体的表达,但抑制乳腺TGF-β的表达。
孕激素和雌激素虽然与催乳素在促进乳腺增生方面具有协同作用,但孕激素和雌激素均拮抗催乳素的生乳作用,因此只有在分娩后孕激素和雌激素水平下降时,催乳素的促进乳汁生成作用才得以表现出来。孕激素的抗催乳素作用是通过抑制催乳素及其受体的表达实现的。分娩后孕激素的水平的急剧下降,导致催乳素的分泌增加,其泌乳作用也因失去孕激素和雌激素的拮抗而得以表现出来。乳糖合成酶复合体成分α-乳汁白蛋白(α-lactoalbumin)与产后的产乳有关,它受催乳素、糖皮质激素的诱导,孕激素则可以拮抗催乳素和糖皮质激素对α-乳白蛋白的诱导作用。除了α-乳汁白蛋白外,孕激素对另一种乳酪蛋白的表达也有抑制作用。
3)孕激素对脑的作用:
雌激素和孕激素参与调控与生殖有关脑区的功能。孕激素受体存在于大脑皮质、下丘脑和垂体。孕激素除了本身对下丘脑和垂体GnRH、FSH和LH的分泌起负反馈调节作用外,还加强雌激素的负反馈作用。
女性基础体温在排卵前出现短暂降低,在排卵后升高0.5℃左右,并在黄体期一直维持在此水平。临床上将这一基础体温的双相变化作为判断排卵的标志之一。妇女在绝经或卵巢摘除后,这种双相的体温变化消失,如果注射孕酮则可以引起基础体温的升高,提示基础体温的升高与孕酮有关。孕酮可能作用于下丘脑的体温调节中枢升高基础体温。
4)孕激素与雌激素的拮抗作用:
孕激素的许多作用是通过拮抗雌激素的作用所实现的,特别是在子宫中的作用尤为突出。当孕激素受体基因敲除后,雌激素对子宫内膜腔上皮和腺上皮的增生作用失去拮抗因素,造成子宫内膜上皮的过度增生。孕激素除了拮抗雌激素对子宫内膜上皮的增生作用外,还在妊娠后期拮抗雌激素对子宫平滑肌的预激作用。因此,妊娠晚期孕激素水平的下降和雌激素水平的升高有利于分娩的启动。孕激素拮抗雌激素作用的机制主要包括以下几个方面:①向下调节子宫的雌激素受体;②拮抗雌激素受体对靶基因的调节作用;③降低雌激素的水平。
2.雌激素
妊娠初期,雌激素主要来源于卵巢黄体。随着胎盘类固醇激素合成功能的成熟,妊娠第7周时,母体循环中胎盘来源的雌激素已经达到50%。以后逐渐增加,妊娠第10周时,切除两侧卵巢已不影响孕妇尿液雌激素的排出量,说明此时雌激素完全来源于胎盘,胎盘已经完全取代卵巢黄体的雌激素合成、分泌功能。胎盘雌激素的分泌量随孕期逐渐增加,足月时达到高峰,分娩后急剧下降(见图5-2)。整个妊娠期由胎盘分泌的雌激素量相当于150个育龄妇女一年的雌激素分泌量,可见胎盘合成和分泌的雌激素量的巨大。胎盘分泌的雌激素主要为雌三醇,此外还有雌酮、17β-雌二醇。妊娠末期尿中雌二醇与雌酮为非孕时黄体期的100倍,而雌三醇为1000倍,且尿中雌激素约90%是雌三醇。
(1)胎盘雌激素的合成
卵巢雌激素合成的经典途径:
灵长类(包括人类)胎盘雌激素的合成途径既不同于体内其他合成雌激素的内分泌腺体,也不同于其他种属动物的胎盘。在卵巢和其他动物的胎盘,雌激素是由胆固醇从头合成的。胆固醇既可以由醋酸盐合成,也可以由LDL摄取。例如,在卵巢内膜细胞内,胆固醇在P-450scc和3β-HSD的作用下,首先合成含有21个碳原子的孕烯醇酮或孕酮,孕烯醇酮和孕酮继续在微粒体细胞色素P-450酶17α-羟化酶(17α-hydroxylase,P-450 C17 )的催化作用下生成雌激素的前体。此过程包括两步:首先孕烯醇酮和孕酮在P-450 C17 的催化下,在C17位发生羟基化,分别形成17-羟基孕烯醇酮和17-羟基孕酮。然后在同一P-450 C17 的催化下,C17位含有两个碳原子的侧链发生裂解,形成含有19个碳原子的雄性激素——脱氢表雄酮和雄烯二酮(C19类固醇)。因此P-450 C17 具有羟化酶和裂解酶的双重催化功能。脱氢表雄酮可在17β-羟基类固醇脱氢酶(17β-HSD)的作用下,C17位上的酮基发生加氢还原成为雄烯二醇。脱氢表雄酮和雄烯二醇又可在3β-HSD的催化下将C3位上的羟基脱氢氧化为酮基和将C5和C6之间的双键转移至C5和C4之间分别形成雄烯二酮和睾丸酮。然后,睾丸酮和雄烯二酮由卵泡内膜细胞(theca cell)转运至卵泡液中,再由卵泡的颗粒细胞(granular cell)摄取,在颗粒细胞中由芳香化酶催化分别生成为雌二醇和雌酮。雌酮还可在17β-HSD的催化下还原为雌二醇。非灵长类动物胎盘雌激素的合成与卵巢类似。
高级灵长类胎盘雌激素合成的非经典途径:
通过卵巢雌激素合成经典途径可以看出,C19类固醇为雌激素合成的前体,17α-羟化酶(P450 c17 )则是转化C21类固醇激素(孕烯醇酮和孕酮)成为C19类固醇(脱氢表雄酮和雄烯二酮)的关键酶。如果组织中存在P450 c17 就可以利用胆固醇或孕激素为雌激素合成提供雄激素前体;如果组织中缺乏P450 c17 则不能利用胆固醇或孕激素为合成雌激素提供雄激素前体。高级灵长类胎盘缺乏P450 c17 ,因此不能利用胆固醇、孕烯醇酮和孕酮为雌激素合成提供雄激素前体。虽然高级灵长类胎盘不能利用孕激素合成雌激素,但高级灵长类胎盘含有高活性的芳香化酶,因此转化C19类固醇(脱氢表雄酮、雄烯二酮、睾丸酮)为雌激素的能力很强。胎盘C19雄激素的来源便成为灵长类胎盘雌激素合成的限速步骤。非灵长类动物胎盘存在17α-羟化酶,因此可利用胆固醇、孕烯醇酮和孕酮合成雌激素。所以,非灵长类动物的妊娠晚期,胎盘P450 c17 在胎儿来源的糖皮质激素的诱导下表达增加,导致孕激素向雌激素的转化增加,因此母体血液孕激素水平下降的同时雌激素水平升高。由于高级灵长类胎盘缺乏P450 c17 ,因此不存在孕激素向雌激素的转化,所以母体血液的孕激素和雌激素水平在妊娠晚期同时升高。
- 上一篇:胎盘内分泌概论
- 下一篇:胎盘C19类固醇的来源
