
两型11β-HSD同工酶
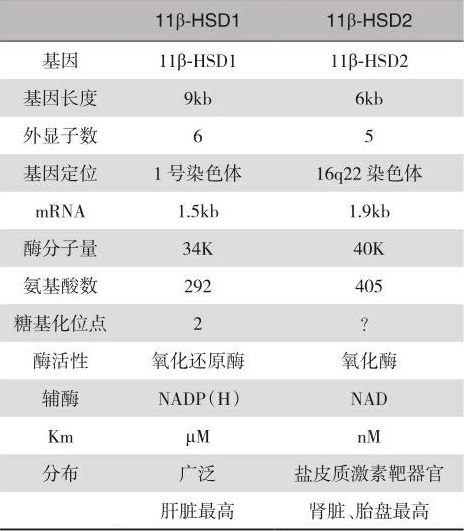
2023-05-05 15:02:10 点击数:随之人们首先从大鼠肝脏分离纯化了11β-HSD,发现此酶主要以糖基化的形式存在于微粒体中,其作用依赖于辅酶NADP(H)。但它并不是单纯的氧化酶,而具有氧化还原酶的双向催化作用,既可使皮质醇脱氢氧化为无活性的17-羟-11-脱氢皮质酮,又可使无活性的17-羟-11-脱氢皮质酮还原为有活性的皮质醇,但以还原酶作用为主。肝脏11β-HSD的催化特性有悖于11β-HSD对盐皮质激素受体的保护作用。进一步的研究还发现,肝脏11β-HSD对皮质醇的米氏常数(Km)在μM水平,高于血液循环糖皮质激素的nM水平,显然也不适应此酶对盐皮质激素受体的保护作用。另外,肝脏11β-HSD及其mRNA也不存在于富含MR受体的肾脏集合小管细胞。更令人难以理解的是AME病人没有肝型11β-HSD基因的变异。以上研究结果均提示,肝型11β-HSD担任保护肾脏MR受体的可能性很小。因此,有人提出肾脏可能还存在一种高亲和力并仅具有脱氢酶单向催化作用的11β-HSD亚型。1992年,Mercer和Krozowski发现,肾脏富含MR的远曲小管和集合管存在一种依赖于辅酶NAD + 的11β-HSD氧化酶。他们把先前发现的依赖于辅酶NADP(H)的肝型11β-HSD称为一型11β-HSD(11β-HSD1),而依赖于辅酶NAD + 的肾型11β-HSD称为二型11β-HSD(11β-HSD2)。1994年,Stewart等对肾脏11β-HSD的酶动力学进行了研究,发现肾脏确实存在一种高亲和力并依赖于辅酶NAD的11β-HSD,它与皮质醇和皮质酮结合的Km值分别为60nM和13nM。1993年,Brown等人首先从人类胎盘部分纯化了11β-HSD2,此酶与已纯化的肝脏11β-HSD相比具有不同的免疫原特性,不为肝脏11β-HSD的抗体所识别,而且具有单向脱氢氧化酶的催化特性。因此人们推测,肾脏11β-HSD的催化特性可能类似胎盘11β-HSD。随后不久人们便分别克隆了羊、人类、兔、大鼠肾脏的11β-HSD2 cRNA,发现11β-HSD2和11β-HSD1是不同的基因产物,11β-HSD2只具有氧化酶的活性,对糖皮质激素的亲和力在nM范围,其mRNA和酶蛋白存在于肾脏的远曲小管和集合小管,AME病人也的确有11β-HSD2基因的变异。从而确认了体内存在两型11β-HSD,11β-HSD2是赋予盐皮质激素受体特异作用的关键因素。表5-1是两型11β-HSD的比较。
表5-1 人类11β-HSD1和2的比较
(2)11β-HSD1和11β-HSD2在胎盘和胎膜中的分布和作用:
糖皮质激素虽然可以促进胎儿肺脏和胃肠道等器官的成熟,但过多的糖皮质激素抑制胎儿的生长发育,并对脑和肾脏产生不良影响,甚至编程胎源性疾病的发生。胚胎初期,胎儿本身基本不产生糖皮质激素。胚胎后期,由于胚胎器官成熟和分娩启动的需要,胎儿肾上腺开始制造自身的糖皮质激素。接近足月时,胎儿血液循环的糖皮质激素75%来源于自身。因此,妊娠期间母体和胎儿之间需要一道屏障以阻挡高于胎儿5~10倍浓度的母体糖皮质激素进入胎儿血液循环。
虽然胎儿的一些器官在胚胎早期就有11β-HSD2的表达,可以灭活进入胎儿体内的糖皮质激素,但胎儿循环中17-羟-11-脱氢皮质酮主要来源于母体,说明糖皮质激素在通过胎盘时,多数已被转化为无活性的17-羟-11-脱氢皮质酮。胎盘11β-HSD2的屏障作用比胎儿本身组织表达的11β-HSD2更为重要。人类胎盘灌流实验则直接表明,灌流入胎盘母体侧绒毛小叶间隙的皮质醇大部分被转化为17-羟-11-脱氢皮质酮并由胎儿侧静脉流出,即糖皮质激素在进入胎儿循环前大部分被胎盘11β-HSD2转化为无活性的17-羟-11-脱氢皮质酮,胎儿组织中11β-HSD2只为避免糖皮质激素作用于胎儿提供了第二道防线。
胎盘是体内11β-HSD2含量最高的器官之一,整个妊娠过程中胎盘11β-HSD的活性都以脱氢氧化酶为主,现已在人类、狒狒、大鼠、羊、猪胎盘中发现有11β-HSD2的存在。免疫组织化学研究发现,胎盘11β-HSD2主要分布于母体血液与胎盘组织交界的胎盘绒毛膜小叶的合体滋养层细胞,使11β-HSD2处于调控母体糖皮质激素进入胎儿循环的前沿。
英国David Barker教授等的流行病学研究表明,相对胎龄低出生体重(胎儿生长受限)并伴有过大胎盘的婴儿,成年后高血压、非胰岛素依赖性糖尿病的发病率较高,此类病人往往死于缺血性心脏病。提示胎儿宫内生长发育的环境可能编程成年疾病即胎源性疾病的发生。
胎儿生长受限产生的原因一般认为与营养不良有关。近年来发现,糖皮质激素同样也为胎儿生长受限发生的重要原因,而且与疾病印迹的关系更为密切。妊娠期营养不良可导致胎盘11β-HSD2的活性降低、胎儿体内糖皮质激素浓度过高。因此,胎盘11β-HSD2的功能异常造成过多糖皮质激素进入胎儿体内可能是胎儿生长受限的重要环节。动物实验发现,大鼠胎盘11β-HSD2的活性与出生体重成正比,与胎盘重量成反比。妊娠期给予大鼠地塞米松或11β-HSD抑制剂CBX,可以导致胎鼠生长受限。此类胎鼠成年后,心血管疾病和糖尿病的发病率均高于正常鼠。人类实验也发现,生长受限胎儿的血液糖皮质激素浓度增加,妊娠期接受糖皮质激素治疗的孕妇,可以导致胎儿生长受限。另外,先天性缺乏11β-HSD2基因的AME病人,出生时也多伴有胎儿生长受限。人类胎盘11β-HSD2的活性虽然与胎盘的重量无相关性,却与出生体重呈正相关。以上动物和人类的观察均表明,胎盘11β-HSD2的功能异常是胎儿生长受限的重要原因。
过去人们普遍认为胎盘只存在11β-HSD2,其实胎盘11β-HSD还原酶的活性在妊娠早期即已存在,而且随妊娠的进展有增加的趋势。免疫组织化学研究表明,11β-HSD1与11β-HSD2的分布截然不同:11β-HSD1存在于人类胎盘的血管内皮、过渡型滋养层细胞、羊膜的上皮细胞和绒毛膜的滋养层细胞及蜕膜细胞,并以绒毛膜的含量为最高。含有11β-HSD2的合体滋养层细胞却不含有11β-HSD1。PCR和Northern分析结果支持免疫组织化学的研究结果,即胎盘、胎膜和蜕膜组织均表达11β-HSD1mRNA,但以绒毛膜的11β-HSD1 mRNA水平为最高。体外培养的绒毛膜滋养层细胞主要表现为11β-HSD1还原酶的活性,而胎盘合体滋养层细胞则主要表现11β-HSD2氧化酶的活性,这也进一步证实了免疫组织化学和mRNA的分析结果。
人类胎盘体外灌流实验发现,将17-羟-11-脱氢皮质酮灌流入胎盘母体侧绒毛小叶间隙,于胎儿侧静脉可以检测到皮质醇,说明胎盘11β-HSD1可能与11β-HSD2相辅相成精细调节糖皮质激素进入胎儿体内的量。胎膜11β-HSD1存在的意义目前尚不清楚,可能参与糖皮质激素的局部作用。实验发现,胎膜11β-HSD1通过转化17-羟-11-脱氢皮质酮为有活性的皮质醇抑制绒毛膜前列腺素降解酶的活性,从而减少前列腺素的降解。胎膜11β-HSD1产生的皮质醇还促进胎膜局部前列腺素的合成。胎膜11β-HSD1的这些作用可能与分娩启动和胎膜的破裂有关。
(二)肽类激素
人类胎盘分泌大量的多肽类激素,发现最早和研究较多的是人绒毛膜促性腺激素(human chorionic gonadotropin,hCG)、人类胎盘生乳素基因家族,包括人类胎盘生乳素(human placental latogen,hPL)和人类变异生长激素(human growth hormone-variant,hGH-V)等,除了这些经典的胎盘肽类激素外,近年来发现胎盘也能够从头合成许多下丘脑、垂体和其他组织分泌的肽类激素,例如促肾上腺皮质激素释放激素(CRH)、促性腺激素释放激素(GnRH)、生长激素释放激素和抑制激素、瘦素、肾素-血管紧张素系统、松弛素、心房钠尿肽和许多脑肠肽等。它们在早期妊娠维持、胎儿生长发育和分娩中起着重要作用。本节介绍几种主要的经典胎盘激素和近年引起关注的胎盘分泌的CRH。
1.人绒毛膜促性腺激素
人绒毛膜促性腺激素(hCG)是胎盘最早能够合成分泌的激素之一,受精后第10天左右即可在母体血液检测到hCG。月经周期的黄体期,卵巢黄体分泌大量的雌激素和孕激素,它们反馈抑制垂体黄体生成素(LH)的分泌。此时如果没有受孕,由于黄体失去了垂体来源的LH的支持,黄体功能只能维持两周左右便萎缩;如果怀孕,胎盘分泌的hCG具有LH的作用,因此可以替代LH维持黄体的功能直到胎盘本身产生孕激素和雌激素的功能成熟。因此hCG是维持早期妊娠的关键激素。
(1)hCG结构:
hCG属于糖蛋白激素家族的一员,此家族还包括LH、FSH和TSH等。这些激素的共同特点是由α和β两个亚基组成,其中α亚基的一级结构相同,但β亚基上的糖基不同;这些激素的β亚基的一级结构和糖基都不相同,β亚基是赋予此家族成员特异功能的亚基。
α亚基由92个氨基酸组成,分子量为14 500,α亚基多肽链中有10个半胱氨酸,它们之间可形成5个二硫键;α亚基多肽链中还含有2个通过N-糖苷键连接于第52位和78位的天冬酰胺的寡聚多糖链。多肽链的糖成分占α亚基分子量的30%。β亚基由145个氨基酸组成,分子量为22 000,分子内有12个半胱氨酸,它们之间可形成6个二硫键。hCGβ亚基与FSH和TSHβ亚基分别存在34%和38%的同源性;hCGβ亚基多肽链的N末端的120个氨基酸与LHβ亚基多肽链存在80%的同源性,而C末端的24个氨基酸为hCG的特异性结构,不存在于LHβ亚基。hCGβ亚基含有6个寡聚多糖链,其中两条链通过N-糖苷键分别与第13和30位的天冬酰胺连接,另外4条多糖链通过O-糖苷键分别与第121、127、132和138位的丝氨酸连接。hCGβ亚基的糖成分占分子量的30%。hCG分子内的二硫键起着稳定hCG三级结构的作用,而多糖成分起着维持hCG与受体结合构象的作用,α亚基和β亚基都是hCG与受体结合所需要的,游离的α亚基和β亚基具有一定的生物活性。当hCG浓度较高时,它还能与TSH受体结合。人类妊娠母体血浆、尿液和胎盘除了存在hCGαβ二聚体外,还存在游离的α亚基、β亚基和β亚基的核心片段(core fragment)。
游离α亚基与hCGαβ二聚体中的α亚基含有的寡聚多糖链不同:游离α亚基N-糖苷键连接的寡聚多糖的分支较多,含有较多的果糖和唾液糖成分;游离α亚基第39位的苏氨酸残基还含有O-糖苷键连接的寡聚多糖。由于游离α亚基的寡聚多糖体积较大,因此妨碍α亚基与β亚基的结合。过去认为游离α亚基没有生物活性,但最近发现它具有一定的生物活性,游离α亚基参与蜕膜催乳素的释放调节。
游离β亚基除了N-糖苷键连接的寡聚多糖与hCG中β亚基稍有不同外,其他结构与hCGβ亚基结构基本一致。只有当游离的β亚基浓度较高时才有一定的生物活性。另外,游离β亚基可以与α亚基结合形成完整的hCG二聚体。
人类胎盘还分泌一种β亚基核心片段,妊娠血清和尿液都含有这种片段,其中尤以尿液含量为高。此片段由hCGβ亚基的第6~40位的氨基酸残基片段通过二硫键与第55~92位氨基酸肽链片段结合而成,分子中含有2个N-糖苷键连接的寡聚多糖链。β亚基核心片段没有生物活性。妊娠血清和尿液中还存在少量结构缺陷的hCGαβ二聚体(nicked αβ dimer),其β亚基缺少第44~52位氨基酸残基。结构缺陷的hCG生物活性只有完整hCG生物活性的20%。
(2)hCG基因结构:
hCGα亚基的基因位于6号染色体上,长度约为9.4kb,包括4个外显子和3个内含子,转录起始位点位于TATAAA共有序列下游第23和25对碱基处,基因启动子位于转录起始位点上游100个碱基以内。α亚基基因启动子部分包括cAMP反应元件(CRE)、滋养层细胞特异性反应元件(TRE)、α-激活因子元件(α-ACT)、连接调节元件(JRE)、糖皮质激素反应元件(GRE)和GnRH调节位点等。cAMP激活蛋白激酶A(PKA),PKA磷酸化转录因子CREB,磷酸化了的CREB形成二聚体,然后与CRE结合并启动hCG α亚基基因的转录。TSE也称上游调节位点(URE),与TSE结合的转录因子目前尚不明了,但TSE和CRE之间可相互作用。JRE可与连接调节因子等几种转录因子结合,JRE可能与hCGα亚基的基础性和特异性表达有关。
hCGβ亚基有6个基因,它们与 LH 基因形成一个基因簇,位于19号染色体上,依次分别为 CGβ7 、 CGβ8 、 CGβ5 、 CGβ1 、 CGβ3 、 LHβ 基因, LHβ 基因过去也称为 CGβ4 。这7个基因的总跨度为50kb,每个基因长度为1.45kb,包含3个外显子和2个内含子。 CGβ3 和 CGβ5 表达的β亚基蛋白相同,由 CGβ7 和 CGβ8 表达的β亚基蛋白与 CGβ3 和 CGβ5 表达的β亚基蛋白有1~2个氨基酸的差别。这4个基因5′末端的结构非常相似,转录起始于翻译启动位点的上游第366个碱基处,转录起始位点周围没有TATA结构。CGβ1和CGβ2的第一个内含子存在变异,它们虽然可以转录mRNA,但目前尚未发现由它们表达的β亚基蛋白。因此,表达hCG β亚基的基因实际上只有4个,分别为 CGβ3 、 CGβ5 、 CGβ7 和 CGβ8 。它们在妊娠早期都有表达,但表达的程度不同,其中 CGβ5 转录的mRNA占hCGβ亚基mRNA总量的65%,由 CGβ7 、 CGβ1 、 CGβ2 表达的mRNA只占hCGβ亚基mRNA总量的2%。另外,hCGβ亚基基因的启动子部分存在多个cAMP反应元件(CRE),提示hCGα和β亚基的基因的转录都可被以cAMP为第二信使的激素所调节。
(3)hCG的分泌:
妊娠6周以前,hCG是由胎盘的细胞滋养层细胞合成和分泌的,第6周以后转为由合体滋养层细胞合成和分泌,此时细胞滋养层细胞只合成少量hCG。hCG合成后通过胞裂外排方式迅速释放入血,细胞内hCG的储存量很少。受精后第7天即可在胚泡中检测到完整分子的hCG,母体血液hCG则出现在月经周期LH高峰后的第7.5~9.5天,此时正是胚泡植入和滋养层细胞开始与母体血液接触的时期,以后母体血液hCG浓度呈指数增加。妊娠第8周时达到高峰(10~15μg/ml),并持续至第12周,然后开始下降,并于第18周以后稳定在较低水平,而且一直维持至妊娠足月,于产后第4天由母体血液中消失(见图5-2)。母体血液hCG水平虽然没有昼夜节律,但存在一定的波动,这主要是滋养层细胞激素分泌的波动特性所决定的。妊娠期尿液hCG的水平与母体血液相平行,如果受孕,妊娠后第6~10周,孕妇尿液的hCG水平可由1IU/ml(妊娠第4周)迅速升高到100IU/ml。因此,根据此现象,临床可以检测尿液hCG的水平判断早期妊娠。胎儿血液hCG浓度的变化规律虽然与母体血液类似,但浓度只有母体血液的3%,提示胎盘hCG主要分泌进入母体血液,少量可以进入胎儿血液。妊娠早期羊水的hCG浓度与血液浓度相仿,但妊娠中、晚期羊水中hCG浓度迅速下降到母体血液水平的20%。羊水hCG除了来源于滋养层细胞外,胎儿肾脏排泄的hCG可能也为羊水hCG的来源。
母体血浆游离α亚基和β亚基水平的变化不同于完整hCG分子。游离α亚基在妊娠第6周时出现在母体血浆,以后逐渐升高,最高时可达500ng/ml,妊娠晚期,游离α亚基可占母体血液α亚基总量的30%~50%。妊娠第3周时母体血浆出现游离β亚基,其变化特征与完整hCG有相似之处,但浓度远远低于hCG,浓度最高时只达到300ng/ml,只占母体血液α亚基总量的3%。游离α亚基和β亚基也存在于羊水,而且占羊水的α和β亚基总量的比例较高,说明完整的hCG主要分泌入母体血液,而分泌入羊水的量比较少。由于β核心片段结合于母体血浆的大分子物质,因此β核心片段的检测易受大分子物质的干扰,妊娠中期母体血液β核心片段的浓度高于妊娠早、晚期。
母体血液hCG的清除呈现快、慢两个时相,半衰期分别为6小时和36小时,20%的hCG经尿液排出,其余部分主要由肾脏的近曲小管、肝脏的网状内皮和卵巢细胞摄取,然后被细胞内的微粒体酶降。母体血液的游离α和β亚基的清除虽然也有两个时相,但清除速度比hCG快得多,半衰期分别为13分钟和40分钟,很少有完整的游离α和β亚基随尿液排出。
(4)合成、分泌的调节:
胎盘hCG的合成和分泌受许多激素、细胞因子和生长因子的调节,但何为调节胎盘hCG分泌的关键因素目前尚不清楚。PKA和PKC的激活都可以磷酸化CREB,CREB通过与hCG基因启动子的CRE结合促进胎盘hCG的合成和释放,而且PKC与PKA通路之间还存在相互作用。
(5)hCG的生物作用:
1)对卵巢功能的影响:
hCG最重要的生物作用是维持卵巢黄体并促进卵巢黄体孕激素和雌激素的合成和释放。排卵后卵巢黄体的形成和功能主要受垂体LH的调节,但随着黄体分泌孕激素和雌激素量的增加,性激素反馈作用于下丘脑和垂体使GnRH和LH的分泌减少。如果此时没有受孕,随着垂体LH的释放减少,黄体功能只能维持2周左右,然后出现萎缩,因此血液循环中的孕激素和雌激素浓度急剧下降,子宫内膜则因为失去了孕激素和雌激素的支持而出现脱落和出血,形成月经。如果受孕,胎盘在妊娠第10天左右开始分泌hCG,并在妊娠第8周达到高峰,此高峰一直持续到妊娠第12周,以后逐渐下降,并于妊娠第18周以后稳定在低水平,直至妊娠足月。hCG具有LH样作用,妊娠初期当垂体来源的LH逐渐减少的情况下,逐渐增多的胎盘hCG取代垂体LH的作用,继续维持卵巢黄体的存在和功能,使卵巢黄体继续分泌孕激素和雌激素,并维持4周左右,直到胎盘本身具有合成孕激素和雌激素的能力。因此,hCG是维持早期妊娠的关键激素。未受孕妇女如果接受hCG治疗可以延长卵巢黄体的寿命、刺激黄体孕激素和雌激素的合成;早期妊娠如果给予孕妇hCG抗体可以导致流产。以上证据表明,hCG为妊娠早期维持黄体功能的主要激素。另外,晚期黄体对hCG的敏感性比早期黄体高。
黄体hCG受体克隆研究发现,此受体对垂体的LH和胎盘的hCG都有很高的亲和力,因此也称为LH/hCG受体。hCG受体为Gs蛋白耦联受体,第二信使为cAMP,cAMP激活的细胞内信号转导途径除了促进黄体细胞LDL受体的表达增加胆固醇的摄取外,还促进P450scc的表达,后者催化胆固醇向孕烯醇酮的转化,最后导致孕激素的合成增加。受孕后在hCG的作用下,妊娠黄体的功能可以维持4周左右,以后虽然血液中的hCG水平仍然较高,但由于黄体对hCG的敏感性降低而出现萎缩。黄体对hCG的失敏原因目前尚不清楚,雌激素可能是其失敏的原因之一。hCG在促进黄体合成孕激素的同时也促进黄体雌激素的合成和释放,雌激素具有抑制hCG释放孕激素的作用,因此随着雌激素的合成增多,hCG对黄体孕激素分泌的影响受到抑制,而hCG促进雌激素的释放作用没有失敏现象。hCG除了促进黄体孕激素和雌激素的分泌外,还促进黄体抑制素和舒缓素的释放,抑制素抑制垂体FSH的释放,从而使妊娠期卵泡的发育受到抑制。
2)对胎盘激素分泌的影响:
hCG除了促进黄体孕激素和雌激素的分泌外,还促进胎盘孕激素的分泌。用hCG抗血清中和胎盘分泌的内源性hCG,胎盘孕激素的分泌减少,提示hCG促进胎盘孕激素的合成和释放,其作用机制类似对黄体的作用。hCG同样促进胎盘雌激素、抑制素、IGF-Ⅱ和PGE 2 的产生。用hCG抗血清可使胎盘滋养层细胞的增生受到抑制,提示内源性的hCG还促进胎盘滋养层的增生。
3)对胎儿类固醇激素合成的影响:
胎儿血液睾丸酮的浓度高峰与胎儿血液和羊水的hCG浓度高峰的时间一致,因此有人提出hCG可能参与胎儿睾丸酮的合成。实验发现,睾丸存在高亲和力的hCG结合位点,hCG可能通过cAMP促进睾丸酮的分泌和睾丸间质细胞的分化。
妊娠15周以前,胎儿肾上腺胎儿带类固醇激素的合成不受垂体ACTH的调控,提示存在调控胎儿肾上腺功能的其他激素。胎儿肾上腺的胎儿带在妊娠第10周左右开始合成脱氢表雄酮,后者为胎盘雌激素合成的前体,此时也恰为血浆hCG高峰出现的时间。因此,hCG很可能为胎儿肾上腺类固醇激素合成的主要调节因素之一。成年人肾上腺的网状带存在hCG受体,虽然相当于网状带的胎儿肾上腺胎儿带是否存在hCG受体目前尚不清楚,但给予新生儿hCG可以使新生儿尿液的hCG排出增加。体外试验也表明,hCG促进胎儿肾上腺脱氢表雄酮的产生。
4)甲状腺:
临床研究发现,患有分泌hCG肿瘤的病人通常有甲状腺亢进的症状,给予正常人hCG也可促进甲状腺的功能。妊娠血浆hCG水平与血浆游离甲状腺激素水平的升高和血浆TSH水平的下降呈一定的相关关系。流产后hCG水平的急剧下降伴随着母体血浆甲状腺激素水平的下降和TSH水平的升高。以上观察说明hCG促进母体甲状腺的功能。动物实验发现,hCG人类妊娠血浆都能促进动物甲状腺的功能,妊娠血浆的作用可被hCG抗血清所中和。这为hCG促进甲状腺功能提供了直接证据。甲状腺细胞膜不仅存在hCG的结合位点,而且hCG和TSH受体还存在交叉反应,说明hCG的促甲状腺功能的作用可以通过甲状腺的hCG受体和TSH受体实现。
2.人类胎盘生乳素
人类胎盘生乳素(human placental lactogen,hPL),又称人类绒毛膜促生长激素(human chorionic somatomammotropin,hCS),为合体滋养层细胞分泌的单链多肽激素,具有生长激素的作用,调节母体与胎儿的物质代谢、促进胎儿生长。最初发现hPL对动物具有很强的催乳作用,故命名为人胎盘催乳素,但后来的研究证明,hPL对人类几乎没有催乳作用,而主要具有促进胎儿生长作用,因此又称为人类绒毛膜促生长激素,但hPL的名称一直沿用至今。
(1)hPL分子结构:
hPL是一个不含糖基的191个氨基酸的单链多肽,分子量为22 279。它与胎盘分泌的变异生长激素、垂体分泌的生长激素和催乳素属于同一家族,hPL结构与生长激素有95%的同源性,与催乳素也有67%的同源性。hPL通过2个二硫键形成与生长激素结构类似的二级、三级结构。hPL与生长激素都可以与催乳素受体结合,而且具有相似的亲和力,进一步的分析发现两者与催乳素受体结合的结构域同源性很高。hPL也可以与生长激素受体结合,但亲和力比生长激素低约2300倍,主要原因是其结合部位的结构不同于生长激素。血浆和胎盘的hPL除了以单体形式存在外,还以二聚体和多聚体的形式存在。多数二聚体是由2个hPL的单链以二硫键首尾结合而成,也有少数二聚体只是hPL单链之间的非共价键结合。
(2) hPL 基因结构:
hPL 基因与生长激素基因共同位于17号染色体上,形成 GH - hPL 基因簇,此基因簇包括转录 hPL 的基因 hCS - A 、 hCS - B 、 hCS - L 和转录正常生长激素的基因 hGH - N 及转录变异生长激素的基因 hGH - V 。这5个基因都是由5个外显子和4个内含子组成,长度约2.0kb,基因的编码区及其两侧的序列结构同源性高达91%~98%。 hCS - A 和 hCS - B 基因为hPL的主要编码基因,由它们表达的hPL前体只在信号肽处有一个氨基酸残基的差别。因此,经过翻译后加工,两者表达的hPL结构完全一致。妊娠早、中、晚期由 hCS - A 基因转录的hPL mRNA各占hPL mRNA总量的60%、60%和85%。 hCS - A 和 hCS - B 基因在其转录起始位点的-30/-25和-85/-80处存在两个共有序列,分别为TATAAA和CATAAA,82%~95%的hPL的转录起始位点位于TATAAA结构下游的第30对碱基处,其余起始于下游第56对碱基处。 hCS - L 基因可以在胎盘转录,但目前尚未在胎盘发现由此基因表达的hPL蛋白。
hCS - A 和 hCS - B 基因的启动子部位存在多种转录因子的反应元件:①GHF-1反应元件:GHF-1为调节垂体生长激素基因表达的转录因子,GHF-1也同样促进胎盘 hCS - A 和 hCS - B 基因的表达,但由于目前尚未发现胎盘有GHF-1蛋白的存在,因此GHF-1在胎盘 hCS - A 和 hCS - B 基因表达调节中的生理意义还有待进一步研究;②Sp-1反应元件: hCS - A 和 hCS - B 基因的启动子部分还存在转录因子Sp-1的结合位点,Sp-1是调节胎盘 hCS - A 和 hCS - B 基因转录的重要转录因子;③TEF-1结合部位:TEF-1为垂体生长激素基因转录作用很强的转录因子,胎盘可能存在类似的转录因子;④甲状腺激素反应元件: hCS - A 和 hCS - B 基因启动子部分还存在多个甲状腺激素反应元件,GHF-1可以影响甲状腺素受体与 hCS - A 和 hCS - B 基因的结合;⑤糖皮质激素受体也可以与 hCS - B 基因结合,但如何调节转录目前尚不清楚;⑥转录抑制因子: hCS - A 和 hCS - B 基因还有两个转录抑制因子的结合部位,分别为PSF-1和PSF-B,PSF-1可以与这两个部位结合,抑制GHF-1启动的hCS-A和hCS-B基因转录。
(3)hPL分布、分泌和代谢:
hPL为胎盘分泌的主要激素之一,胎盘每天分泌约0.3~1.0g的hPL,足月时hPLmRNA占胎盘总量的5%。免疫组织化学和原位杂交方法都证明,妊娠前6个周,hPL主要由细胞滋养层所合成、分泌,以后随着细胞滋养层细胞向合体滋养层的融合、分化,合体滋养层细胞成为hPL的主要合成细胞。
妊娠第3周左右就可在胎盘和母体血浆检测到hPL,母体血浆hPL水平在妊娠早期呈指数性急剧升高,妊娠中期以后增加速度缓慢,直至妊娠足月(见图5-2)。足月时母体血浆hPL的浓度为5~15μg/ml,其中二聚体和多聚体的比例少于10%,少量的hPL与巨球蛋白相结合。母体血液循环的hPL的半衰期为10~20分钟,由尿液排出的hPL主要以代谢产物为主,少量为完整的hPL。胎儿血液循环中hPL的浓度低于母体循环,妊娠12~20周时胎儿循环hPL的浓度为400~500ng/ ml,足月时下降为20~30ng/ml。胎盘生乳素还可分泌进入羊水,妊娠第11周时羊水hPL浓度就已达到400ng/ml,以后不断升高,妊娠中期开始下降,足月时降至妊娠早期水平。胎盘生乳素进入胎儿循环和羊水的途径目前尚不清楚。
(4)胎盘hPL合成和分泌的调节:
胎盘首先合成带有信号肽的hPL前体,此前体为217个氨基酸,经翻译后加工脱掉号肽,成为成熟的hPL。hPL的翻译后加工和分泌过程为经典的蛋白/肽类激素途径。影响hPL合成和分泌的因素很多,其中包括胎盘体积和合成hPL的细胞数目增加、能量代谢产物、多种激素和生长因子等,但影响hPL表达的主要调节因素目前并未阐明。
(5)hPL的生物作用:
由于hPL的结构与催乳素和生长激素类似,hPL与催乳素和生长激素受体又存在交叉结合,因此hPL的许多功能与催乳素和生长激素的功能类似,所以hPL又称为人类绒毛膜生长催乳激素(human chorionic somatomammotropin,hCS)。生长激素和催乳素受体属于造血因子类受体家族。此类受体细胞内部分虽然不具有酪氨酸蛋白激酶的活性,但与其结合的蛋白质具有酪氨酸蛋白激酶的活性,与此类受体结合的常见蛋白质为JAK酪氨酸蛋白激酶。当配基与受体结合后,导致JAK和受体的磷酸化,磷酸化的JAK进一步对细胞内的其他与生长相关的蛋白质进行磷酸化,磷酸化的蛋白质转位到细胞核内,与其他转录因子一道调节靶基因的转录。胎盘生乳素的催乳作用在动物身上比较明显,对于人体不明显。在人类,hPL的主要作用为促进细胞的增生、影响能量代谢和保证胎儿对营养物质的需求等。
1)对能量代谢的影响:
妊娠中、后期母体的糖代谢和脂类代谢发生了一些适应性变化,以保证胎儿对葡萄糖的需求。这些变化包括胰岛素的基础分泌和葡萄糖诱导分泌增加、某些组织对胰岛素敏感性降低、糖耐受力异常和脂类代谢增加等,hPL是造成母体产生这些适应性代谢变化的主要激素之一。
hPL对胰岛β细胞的胰岛素的表达、分泌具有直接的促进作用。妊娠时母体血液胰岛素水平增加可能与hPL有关。hPL造成的某些组织对胰岛素的失敏可以降低母体组织对葡萄糖的利用,这样可以保证胎儿对营养物质的需要。葡萄糖耐受实验表明,hPL可以使母体葡萄糖的耐受力降低,给予孕妇葡萄糖可以导致血液葡萄糖浓度异常升高。hPL还促进母体脂肪的水解,给予hPL可以导致血液游离脂肪酸、酮体和甘油水平的升高。在能量充足的状态下,hPL可以增加脂肪细胞葡萄糖的摄取、促进脂肪酸重新脂化为甘油三酯以贮存能量,以便饥饿时可以动员更多的母体脂肪供给母体能量,这样可以节省葡萄糖以保证胎儿的需求。
值得指出的是,hPL尽管可以与生长激素受体结合,但亲和力比生长激素低2300多倍,而胎盘产生的变异生长激素与生长激素受体的亲和力却很高,变异生长激素与催乳素受体亲和力却比较低。因此,妊娠时胎盘分泌的hPL和变异生长激素虽然都影响能量代谢,但侧重面不同。hPL具有促进胰岛细胞胰岛素表达的作用但变异生长激素无此作用;变异生长激素具有较强的脂肪水解作用,而hPL的这方面作用则比较弱。
2)对胎儿生长的影响:
过去认为hPL的促生长作用只有生长激素的1%,因此hPL本身对胎儿生长的作用甚微,hPL主要通过影响母体的能量代谢间接影响胎儿的发育。但现在发现胎儿的许多器官组织表达hPL受体,如肝脏、骨骼肌、皮肤、肾上腺、心脏、小肠、肾脏和脑组织等。体外试验发现,hPL促进胎儿成纤维细胞、肝细胞、肌细胞对氨基酸、胸腺嘧啶脱氧核苷的摄取,促进胎儿组织胰岛素样生长因子及其结合蛋白的分泌,hPL产生这些作用所需的浓度与妊娠中期胎儿体内的hPL浓度类似。
3)对乳腺功能的影响:
妊娠期乳腺的发育包括乳腺导管、乳腺小泡的增生和乳汁蛋白的表达等。hPL对动物乳腺具有很明显的促泌乳作用,但对人类乳腺的泌乳作用不很明显。实验发现,hPL可以促进乳腺肿瘤的增殖和乳腺导管上皮的DNA合成。因此,hPL对人类乳腺的主要作用为促进细胞的增生,对乳腺分泌的影响还有待进一步研究。
4)对卵巢功能的影响:
先天性 hCS - A 和 hCS - B 基因缺陷的孕妇血液hPL和变异生长激素浓度很低,甚至缺如。但在这种情况下,妊娠仍能正常进行,而且母体血液的雌激素、孕激素、hCG、PRL和GH的浓度正常,产后泌乳、胎儿出生时和出生后的发育也正常,说明hPL在妊娠中的作用并不关键。但最近有一 hPL 基因缺陷病例报道,hPL基因缺陷时虽然妊娠可以维持,但胎儿出生时发育迟缓,妊娠期母体有轻微的先兆子痫症状。有人认为,hPL很可能是妊娠期垂体催乳素和生长激素的后备军,一般情况下,垂体激素的作用已经足够满足妊娠的需要,但在饥饿状态下,需要大量的生长激素才能保证胎儿能量的需求,这时由于胎盘分泌的hPL与生长激素和催乳素的作用交叉,因此可以弥补生长激素的分泌不足。
3.人类变异生长激素
人类变异生长激素(human growth hormone variant,hGH-V)为胎盘分泌的生长激素,与hPL同属生长激素家族。hGH-V的偶然发现与妊娠期生长激素的测定有关,当人们用两种生长激素的单克隆抗体测定妊娠血浆生长激素的变化特征时,发现用一种抗体测定的生长激素水平在妊娠25周后明显上升,而另一种抗体测得的生长激素水平反而下降,用第一种抗体也可以在胎盘检测到生长激素的存在,但第二种抗体不能检测到胎盘生长激素。说明妊娠25周后,母体血浆生长激素的增加来源于胎盘分泌的一种抗原性与垂体生长激素有交叉的生长激素。随着胎盘生长激素的纯化、cDNA的克隆,人们认识到胎盘生长激素的结构不同于垂体生长激素,是一种变异的生长激素。
(1)胎盘hGH-V及其基因的结构:
hGH-V与垂体分泌的生长激素hGH-N一样也是含有191个氨基酸的多肽,其分子内存在2个二硫键,分别将Cys53和Cys165、Cys 182和Cys189连接。分子内第140位的天门冬酰胺发生糖基化。hGH-V前体结构与hGH前体相比存在15个氨基酸的差别,前体经翻译后加工脱掉信号肽成为成熟激素后,两者之间还有13个氨基酸的差别。虽然hGH-V、hPL和hGH都能与催乳素受体结合,但hGH-V与催乳素受体的结合部位结构不同于hPL和hGH,因此结合机制也不相同,hGH-V与催乳素受体的结合机制更接近于催乳素。hGH-V基因转录产物经过不同剪切还可以形成另外一种分子形式的mRNA,理论上讲此mRNA可以翻译出另外一种hGH-V或hGH-V2。根据推测hGHV2应含有230个氨基酸,其N末端的126个氨基酸残基与hGH-V相同,但C末端区别较大,而且不含糖基化位点,二硫键的形成位置也与经典的hGH-V不一样,因此其三级结构也不同于hCG-V和hGH。由于hGH-V2分子内存在疏水性结构,因此有人认为hGH-V2可能是细胞膜蛋白。
hGH - V 的基因属于 GH - PL 基因簇的一员,也由5个外显子组成。如上所述, hGH - V 基因转录的mRNA,经不同剪切后可以产生两种mRNA,分别表达 hGH - V 和 hGH - V2 。 hGH - V2 mRNA只占 hGH - V mRNA总量的5%~15%。 hGH - V 基因的表达也不同于 hGH - N ,后者外显子3的前15个密码不被转录。 hGH - V 基因的转录起始位点位于TATAAA序列的下游第30个碱基处,其启动子部分可以与转录因子GHF-1结合,后者是很强的 hCG 基因转录激活因子。抑制子PSF-1也可以与 hGH - V 基因结合,抑制 GHF - 1 的转录激活作用。由于垂体表达大量的PSF-1,因此垂体 hGH - V 基因不能表达,而只能表达 hGH - N 。
(2)胎盘hGH-V的分布与分泌:
hGH-V与hPL的基因和蛋白结构类似,因此为研究hGH-V的特异性分布带来了困难。用针对 hGH - V2 基因内含子4的探针进行原位杂交,发现 hGH - V2 基因存在于胎盘合体滋养层细胞,因此推测 hGH - V 基因也存在于此型细胞,但目前尚无这方面的直接证据。用与垂体GH和hGH-V有交叉反应的单克隆抗体对胎盘进行免疫组织化学染色,发现hGH-V蛋白存在于胎盘的合体滋养层细胞和胎盘基底板的细胞,但此抗体与hGH-V2的交叉反应尚不清楚。母体血浆hGH-V的测定也存在抗体特异性问题,目前关于母体血浆hGH-V水平的数据主要通过两种特异性不同的单克隆抗体所获得的,一种抗体只能识别hGH-N,另一种既可识别hGH-N,又可识别hGH-V。妊娠21~26周时用两种抗体测定的母体血浆生长激素样免疫活性物质开始出现分离,与hGH-V有交叉反应的单克隆抗体测得的血浆GH水平在妊娠21~26周时开始升高,并持续到妊娠第36周,以后稳定在这个水平,直到足月妊娠。可以认为以上变化特点反映了妊娠hGH-V分泌的特点。由于妊娠第9周时,胎盘就可检测到hGH-V和hGH-V2mRNA,因此有人认为胎盘hGH-V的分泌可能早于妊娠21周,只是受测定方法灵敏度所限未能测出。足月羊水和胎儿血液中也未能检测到hGH-V。母体血液hGH-V水平不同于血液的垂体生长激素水平,hGH-V水平没有昼夜节律。甲状腺激素促进胎盘hGH-V基因的表达,调节垂体生长激素表达和分泌的下丘脑激素-生长激素释放激素(GHRH)对胎盘hGH-V的分泌没有影响。
(3)胎盘hGH-V的生物作用:
胎盘hGH-V的生物作用目前尚不明了。在大鼠hGH-V具有生长激素和催乳素的双重作用,它既可以促进大鼠淋巴肿瘤细胞的分裂,又可与大鼠肝脏的催乳素受体结合,但与催乳素受体的亲和力和促细胞分裂作用均远远弱于垂体生长激素。胎盘hGH-V可以促进切除垂体大鼠的体重增长、葡萄糖氧化和脂肪组织的脂肪水解,其作用强度类似垂体生长激素。在人类胎盘,hGH-V是否具有同样的作用目前尚无实验证据。
胎盘细胞膜存在hGH-V的受体,因此hGH-V在胎盘局部可能有旁分泌和自分泌作用。在人类,hGH-V可能与IGF-Ⅰ的表达有关,两者在妊娠中的变化呈一定的相关关系。由于胎盘hGH-V与生长激素受体有交叉结合,因此hGH-V在人类妊娠中的促躯体生长作用可能类似于生长激素。虽然过去认为 hGH - V 基因的先天性缺陷不影响妊娠的正常进行,但最新的临床观察发现, hGH - V 和 hPL 基因的先天性缺陷可以导致胎儿生长受限,母体发生轻微先兆性子痫。
4.促肾上腺皮质激素释放激素
促肾上腺皮质激素释放激素(corticotrophin releasinghormone,CRH)为下丘脑分泌的41肽激素。CRH的主要作用为促进垂体前叶阿黑皮素原(pro-opiomelanocortin,POMC)来源激素的合成与释放。阿黑皮素原经酶分解出促肾上腺皮质激素(ACTH)、β-趋脂素(β-LPH)和β-内啡肽(β-EP)。CRH主要由下丘脑的室旁核小细胞神经元分泌,经正中隆起的垂体门脉系统转运至垂体前叶,调节ACTH的释放。CRH还广泛分布于下丘脑以外的中枢其他部位,如大脑皮层、小脑、丘脑、海马和脑干等。肾上腺、胃肠道、肺等外周组织也表达不等量的CRH。近年来发现胎盘胎膜分泌大量的CRH,胎盘成为妊娠期CRH的主要分泌器官。
- 上一篇:对胎盘孕激素合成的调节作用
- 下一篇:妊娠期母体血液CRH水平:
